pISSN : 3058-423X eISSN: 3058-4302
Open Access, Peer-reviewed
pISSN : 3058-423X eISSN: 3058-4302
Open Access, Peer-reviewed

Kanon Tsurumi,Maki Kakurai,Kazutoshi Harada,Toshio Demitsu,Akira Watanabe,Koichi Makimura,Rui Kano
10.17966/JMI.2026.31.2.70 Epub 2026 June 30
Abstract
Background: Dermatophytosis caused by Microsporum canis infection is a common cutaneous mycosis in small animals. As a zoonotic disease in humans, M. canis infection causes highly inflammatory tinea cruris or tinea corporis, which are treated with oral terbinafine (TBF) or itraconazole (ITCZ) and/or topical antifungal agents. In recent years, dermatophyte resistance to antifungal drugs has gained clinical importance as it impedes dermatophytosis treatment.
Objective: The mechanism of azole ITCZ tolerance in the dermatophyte M. canis was investigated through transcriptome analysis of the ITCZ-tolerant strain DTMU25-001.
Methods: Strain DTMU25-001 was cultured with and without ITCZ, after which total RNA was extracted from mycelial samples and sequenced using a DNBSEQ-G400 system.
Results: A total of 8,849 expressed genes were detected in samples of both RNAs, and gene expression levels were compared between the ITCZ-treated and ITCZ-free samples. Treatment with ITCZ upregulated the expression of 24 genes >2-fold over the untreated sample. Heat shock protein and ABC transporter genes were the most highly expressed and were presumed to be related to the tolerance mechanism. The expression levels of 79 identified genes decreased approximately ≤0.5-fold with the addition of ITCZ.
Conclusion: ITCZ addition reduced the expression of many genes in M. canis cells, which could be related to ITCZ tolerance of this organism.
Keywords
Gene expression Itraconazole Microsporum canis Transcriptome analysis
Dermatophytosis caused by Microsporum canis infection is a common cutaneous mycosis in small animals. As a zoonotic disease in humans1,2, M. canis infection causes highly inflammatory tinea cruris or tinea corporis, which are treated with oral terbinafine (TBF) or itraconazole (ITCZ) and/ or topical antifungal agents. In recent years, dermatophyte resistance to antifungal drugs has emerged, impeding der- matophytosis treatment3,4.
We have previously described the initial isolation of a TBF-resistant M. canis strain from a case of feline dermatophytosis in China in 20185. The isolated strain exhibited a TBF minimum inhibitory concentration (MIC) of >32 μg/mL but remained susceptible to ITCZ (MIC: 0.023 μg/mL)5,6. We later reported the first isolation of an ITCZ-tolerant M. canis strain from a case of human tinea corporis that was resistant to oral ITCZ therapy7. The isolate was ITCZ-tolerant (MIC: 1 μg/mL) and susceptible to TBF (MIC: <0.03 μg/mL), ravuconazole (RVCZ; MIC: 0.06 μg/mL), and luliconazole (LLCZ; MIC: <0.03 μg/ mL)7. As the patient had a pet cat diagnosed with dermato- phytosis, the infection was probably acquired from the cat7. Such cases have raised concerns that the incidence of pet-acquired infections with ITCZ-tolerant dermatophytes is in- creasing. However, the mechanism of ITCZ tolerance in zoophilic M. canis is not as clearly understood as in anthro- pophilic dermatophytes like Trichophyton interdigitale and T. rubrum8. In this study, as a One Health approach, we per- formed a transcriptome analysis to elucidate the tolerance mechanism of ITCZ-tolerant M. canis isolated from a case of human dermatophytosis.
1. Strain and antifungal susceptibility testing
Strain DTMU25-001 of M. canis was isolated from a patient with tinea faciei and corporis whose pet had a case of feline dermatophytosis in 20257.
After determining the MICs for these agents9,10, the cultures were resuspended, and a 10-μL aliquot of the suspension in each well was inoculated onto potato dextrose agar (Kanoto Chemical Co., Inc., Tokyo, Japan) plates using a pipette. After 7-10 days of incubation at 28℃, the minimum fungicidal concentration (MFC) was determined as the lowest con- centration of drug at which colony growth was observed11.
2. Next-generation sequence analysis of whole-RNA samples
Strain DTMU25-001 was cultured in Sabouraud's dextrose broth (SDB: 1% peptone and 2% glucose) at 28℃ with shaking for 5 days. Culture medium containing mycelial cells was incubated with and without ITCZ at a concentration of 0.2 μg/mL for an additional 3 h at 28℃. Mycelia samples (approximately 100-200 mg) were flash-frozen in liquid nitrogen and ground to obtain a homogeneous powder. Total RNA was extracted using the RNeasy Total RNA kit (QIAGEN, Tokyo, Japan). Approximately 1 μg/μL of total RNA from ITCZ-treated and ITCZ-free cultures was stored at -80℃ until analysis.
The mRNA enrichment samples were sequenced using oligodT from whole RNA (300 ng of RNA) using a DNBSEQ-G400 system (MGI Tech Co., Ltd. Tokyo, Japan), and the resulting data were analyzed by Bioengineering Lab. Co., Ltd. (Kanagawa, Japan; https://gikenbio.com). Complementary DNA (cDNA) libraries were synthesized using an MGIEasy Fast RNA Library Prep Set (MGI Tech) and transformed into circular libraries using an MGIEasy Dual Barcode Circularization kit (MGI Tech) for sequencing analysis. For next-generation sequencing analysis of the cyclic cDNA libraries, a DNBSEQ-G400RS High-Throughput Sequencing kit (MGI Tech) and High-Throughput Paired-End Sequencing Primer kit (App-D; MGI Tech) were used to synthesize samples for DNA nano- balls. Whole-RNA sequencing (MGI DNBSEQ-G400RS) and data analysis of the two samples were performed by Bio- engineering Lab. Co., Ltd. (Kanagawa, Japan).
3. Real-time quantitative polymerase chain reaction analysis of heat shock protein and ABC transporter genes
The expression levels of heat shock protein and ABC trans- porter genes were analyzed using real-time quantitative polymerase chain reaction (RT-qPCR) as follows. First, cDNA samples synthesized from 0.5 μg of RNA were amplified using RT-qPCR (Thermal Cycler Dice; Takara) and analyzed as described in the previous report6.
The following primer pair was used to amplify fragments of the open reading frames of target genes: heat shock protein gene (293-bp fragment), forward primer 5'-GCCCT- TGGGCTAAACAATGCCGT-3' and reverse primer 5'-GTATAG- AGCCAACGAGAGTGAGGGCG-3', corresponding to nucleo- tides 56-78 and 339-358, respectively, of the M. canis heat shock protein gene (M. canis CBS 113480 heat shock pro- tein [MCYG_07264], partial mRNA, GenBank accession no. XM_002843435.1).
The following primer pair was used to analyze the ABC transporter gene (302-bp fragment): forward primer 5'-GGAACAACAGCCTTCAATCCACC-3' and reverse primer 5'-CACCGGGAATTTCTCCGTCG-3', corresponding to nucleo- tides 47-69 and 315-340, respectively, of the M. canis ABC transporter gene (M. canis CBS 113480 ABC transporter [MCYG_02928], partial mRNA, GenBank accession no. XM_002847376).
RT-qPCR assays and data analysis (ΔΔCT method) for relative quantitation were conducted using Thermal Cycler Dice Real-Time System software, version 5.11C (Takara). Basal expres- sion levels were estimated based on normalization to the level of actin-encoding transcripts in RNA from cultures under the assumption that an equivalent total RNA input and observed equal PCR efficiency would provide comparable threshold cycle (Ct) values. Gene expression levels were normalized to that of the actin-encoding gene, and values were compared between strains cultured in SDB with and without 0.2 μg/mL ITCZ. All RT-qPCR experiments were per- formed in triplicate. To identify any significant differences in the data, we performed a Student's nonpair t-test with Excel (Microsoft® Excel® 2021).
1. MFCs for M. canis strain DTMU25-001
The MFCs for strain DTMU25-001 were 0.25 μg/mL for TBF, >32 μg/mL for ITCZ, 2 μg/mL for RVCZ, and <0.03 μg/ mL for LLCZ.
2. Comparison of gene expression under ITCZ-treatment and ITCZ-free conditions
Approximately 7,500 Mb were sequenced for both samples, with approximately 24 million paired reads for each (Sup- plemental Fig. 1 and 2). After annotation, the data were mapped to M. canis CBS 113480 (assembly ASM15114v1), which is registered in GenBank, confirming approximately 98% identity between the RNA sequences of both samples.
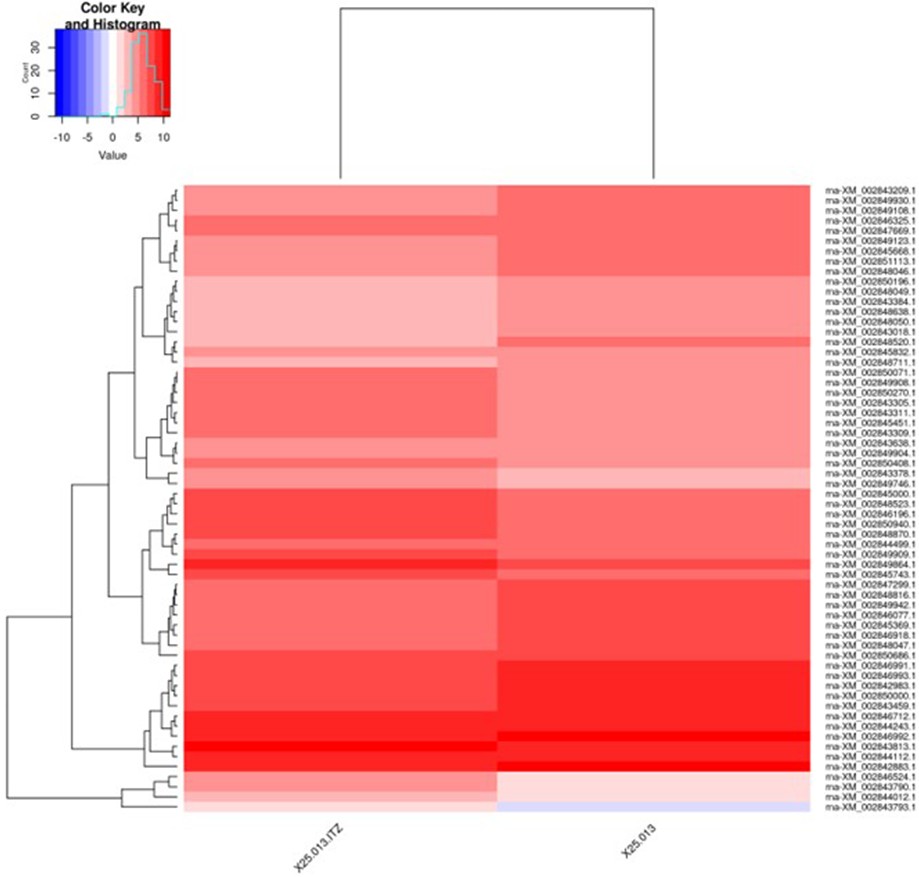
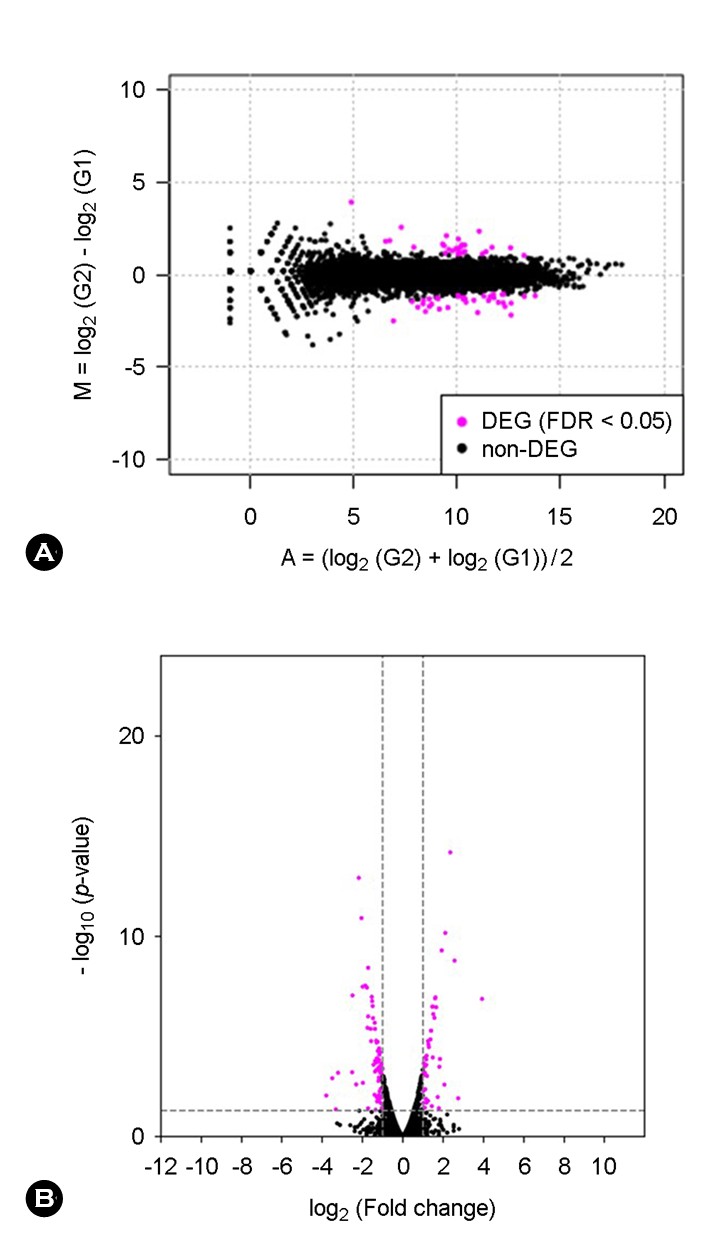
A total of 8,849 expressed genes were detected in both sample RNAs, and the expression levels were compared between the ITCZ-treated and ITCZ-free samples. Among the compared genes, those for which the expression changed by a factor of >2 or ≤0.5 between ITCZ addition and no addition were plotted from highest to lowest expression level (Supplemental Tables 1 and 2). For 24 genes, the ex- pression increased >2-fold following treatment with ITCZ, with the heat shock protein gene (GenBank accession no. XM_002843435.1) and ABC transporter gene (GenBank accession no. XM_002847376.1) being the most highly ex- pressed, and thus presumed to be involved in tolerance (Supplemental Table 1). By comparison, the expression of 79 genes declined approximately ≤0.5-fold after the addition of ITCZ (Supplemental Table 2).
|
Accession
number |
Length |
|
Lead count |
|
TPM (Transcripts Per
Million) |
|
Gene names registered |
||
|
ITZ-Added |
Control |
ITZ-Added |
Control |
||||||
|
XM_002843435.1 |
807 |
|
255,536 |
143,590 |
|
16727.03023 |
8626.06212 |
|
Heat shock protein |
|
XM_002847376.1 |
4,419 |
|
79,633 |
24,150 |
|
951.9395257 |
264.9445889 |
|
ABC transporter |
|
XM_002847375.1 |
23,745 |
|
65,488 |
17,447 |
|
145.6900316 |
35.62136092 |
|
Nonribosomal peptide |
|
XM_002849340.1 |
1,029 |
|
63,545 |
34,508 |
|
3262.167775 |
1625.797065 |
|
Alcohol dehydrogenase |
|
XM_002845285.1 |
2,580 |
|
38,085 |
12,838 |
|
779.7843917 |
241.2345932 |
|
Conserved hypothetical |
|
XM_002844211.1 |
342 |
|
32,092 |
12,201 |
|
4956.909176 |
1729.542513 |
|
Conserved hypothetical |
|
XM_002850512.1 |
1,230 |
|
31,895 |
15,912 |
|
1369.801947 |
627.1646613 |
|
Mannitol-1-phosphate |
|
XM_002848719.1 |
2,088 |
|
27,119 |
2,426 |
|
686.0937463 |
56.32772652 |
|
Peptidase S8 and S53 |
|
XM_002846389.1 |
2,199 |
|
24,105 |
5,948 |
|
579.058166 |
131.1316878 |
|
Conserved hypothetical |
|
XM_002850536.1 |
750 |
|
21,605 |
4,463 |
|
1521.714894 |
288.4878613 |
|
Macrophage binding |
|
XM_002847443.1 |
2,694 |
|
15,123 |
2,441 |
|
296.5381985 |
43.9270571 |
|
C6 transcription factor
FacB |
|
XM_002847470.1 |
1,683 |
|
12,816 |
4,006 |
|
402.2616835 |
115.3954671 |
|
Glutamate decarboxylase |
|
XM_002844579.1 |
1,062 |
|
5,801 |
686 |
|
288.5483749 |
31.31565642 |
|
Glyoxylate reductase |
|
XM_002847067.1 |
3,195 |
|
4,585 |
1,611 |
|
75.80690425 |
24.4448062 |
|
Fatty acid hydroxylase |
|
XM_002848777.1 |
1,080 |
|
4,335 |
1,047 |
|
212.0340606 |
46.99859193 |
|
PQ loop repeat protein |
|
XM_002843246.1 |
543 |
|
2,984 |
987 |
|
290.2948112 |
88.12096485 |
|
Conserved hypothetical |
|
XM_002844128.1 |
474 |
|
2,738 |
454 |
|
305.1373612 |
46.43435654 |
|
Conserved hypothetical |
|
XM_002845967.1 |
843 |
|
2,445 |
828 |
|
153.2115735 |
47.61729083 |
|
Aldo-keto reductase |
|
XM_002849064.1 |
1,245 |
|
2,310 |
546 |
|
98.01282626 |
21.26107463 |
|
Sulfite oxidase |
|
XM_002843813.1 |
610 |
|
2,200 |
594 |
|
190.5167349 |
47.20831838 |
|
Uncharacterized protein |
|
XM_002845648.1 |
606 |
|
2,175 |
905 |
|
189.595018 |
72.39988488 |
|
Conserved hypothetical |
|
XM_002846339.1 |
1,515 |
|
1,989 |
172 |
|
69.35255003 |
5.503991248 |
|
6-phosphogluconate |
|
XM_002845426.1 |
1,167 |
|
1,927 |
319 |
|
87.2270415 |
13.25200978 |
|
NAD binding |
|
XM_002845443.1 |
864 |
|
1,913 |
493 |
|
116.9611182 |
27.66273379 |
|
Conserved hypothetical |
|
Accession
number |
Length |
|
Lead count |
|
TPM (Transcripts Per
Million) |
|
Gene names registered |
||
|
ITZ-Added |
Control |
ITZ-Added |
Control |
||||||
|
XM_002846943.1 |
2,727 |
|
1,574 |
3,171 |
|
30.49017199 |
56.37324369 |
|
GTP-binding protein 2 |
|
XM_002847168.1 |
438 |
|
1,438 |
2,887 |
|
173.4303336 |
319.5468891 |
|
Conserved hypothetical |
|
XM_002847348.1 |
1,425 |
|
1,241 |
2,741 |
|
46.00417017 |
93.25155698 |
|
Zinc finger protein zpr1 |
|
XM_002845039.1 |
1,674 |
|
1,231 |
2,459 |
|
38.84569387 |
71.21393694 |
|
Phosphotransferase |
|
XM_002847378.1 |
958 |
|
1,149 |
2,337 |
|
63.35702899 |
118.2646971 |
|
Vegetative cell wall |
|
XM_002847978.1 |
747 |
|
1,003 |
6,147 |
|
70.92847383 |
398.937197 |
|
Glutathione transferase |
|
XM_002851171.1 |
1,689 |
|
915 |
3,243 |
|
28.61750244 |
93.08489639 |
|
Asparagine synthetase 1 |
|
XM_002850374.1 |
1,203 |
|
817 |
2,289 |
|
35.8753968 |
92.24484085 |
|
4-hydroxyphenylpyruvate |
|
XM_002843209.1 |
498 |
|
816 |
2,069 |
|
86.55678163 |
201.4155833 |
|
YjgH family protein |
|
XM_002845511.1 |
963 |
|
704 |
3,588 |
|
38.61772238 |
180.6292455 |
|
Uncharacterized protein |
|
XM_002842671.1 |
1,170 |
|
686 |
1,724 |
|
30.97266235 |
71.43537359 |
|
2-oxoisovalerate |
|
XM_002849403.1 |
2,379 |
|
685 |
1,379 |
|
15.21025214 |
28.1016451 |
|
Dipeptidylpeptidase 4 |
|
XM_002842689.1 |
7,755 |
|
680 |
1,839 |
|
4.631987669 |
11.49639951 |
|
Polyketide synthase |
|
XM_002843215.1 |
1,753 |
|
668 |
1,630 |
|
20.12958542 |
45.07830824 |
|
Uncharacterized protein |
|
XM_002846325.1 |
1,191 |
|
668 |
3,137 |
|
29.62818072 |
127.6922907 |
|
Conserved hypothetical |
|
XM_002842791.1 |
1,011 |
|
645 |
1,485 |
|
33.70146989 |
71.20938232 |
|
3-hydroxyisobutyrate |
|
XM_002844656.1 |
1,296 |
|
642 |
1,754 |
|
26.16798671 |
65.61248826 |
|
Isovaleryl-CoA |
|
XM_002847159.1 |
1,095 |
|
637 |
1,337 |
|
30.73021489 |
59.19420724 |
|
Conserved hypothetical |
|
XM_002844577.1 |
960 |
|
609 |
1,240 |
|
33.51091943 |
62.61990043 |
|
Hydroxyquinol |
|
XM_002843709.1 |
921 |
|
607 |
1,554 |
|
34.81523612 |
81.80000022 |
|
Polysaccharide deacetylase |
|
XM_002844989.1 |
1,845 |
|
601 |
1,613 |
|
17.20752406 |
42.38380252 |
|
3'-phosphate cyclase |
|
XM_002843038.1 |
1,503 |
|
597 |
1,427 |
|
20.98242283 |
46.02850964 |
|
FAD binding
domain-containing protein |
|
XM_002846909.1 |
1,344 |
|
577 |
2,776 |
|
22.67863067 |
100.1341265 |
|
Translation initiation |
|
XM_002850271.1 |
726 |
|
576 |
2,630 |
|
41.91081891 |
175.6228612 |
|
Acetyltransferase |
|
XM_002843623.1 |
1,728 |
|
575 |
2,665 |
|
17.57779481 |
74.76793667 |
|
Conserved hypothetical |
|
XM_002847902.1 |
537 |
|
569 |
1,349 |
|
55.97295878 |
121.786622 |
|
Conserved hypothetical |
|
XM_002847261.1 |
1,398 |
|
566 |
2,885 |
|
21.38698397 |
100.0461928 |
|
Conserved hypothetical |
|
XM_002850779.1 |
330 |
|
549 |
1,410 |
|
87.88174841 |
207.1414888 |
|
Uncharacterized protein |
|
XM_002851237.1 |
1,830 |
|
526 |
1,039 |
|
15.18360645 |
27.52493984 |
|
MPE1 |
|
XM_002846799.1 |
867 |
|
520 |
1,805 |
|
31.68287108 |
100.9299433 |
|
Conserved hypothetical |
|
XM_002843202.1 |
2,007 |
|
516 |
1,337 |
|
13.58133974 |
32.29579319 |
|
Activating signal |
|
XM_002843911.1 |
1,038 |
|
515 |
1,087 |
|
26.20898242 |
50.76847419 |
|
Uncharacterized protein |
|
XM_002847252.1 |
1,848 |
|
514 |
2,006 |
|
14.69269408 |
52.62485139 |
|
Methionyl-tRNA |
|
XM_002842694.1 |
1,761 |
|
497 |
1,666 |
|
14.90861559 |
45.86459487 |
|
Laccase-1 |
|
XM_002850980.1 |
1,617 |
|
489 |
1,106 |
|
15.97493587 |
33.15942779 |
|
Esterase |
|
XM_002844658.1 |
2,139 |
|
487 |
2,035 |
|
12.02703184 |
46.12278781 |
|
Methylcrotonoyl-CoA |
|
XM_002846163.1 |
996 |
|
480 |
1,272 |
|
25.45787695 |
61.91411842 |
|
Conserved hypothetical |
|
XM_002849679.1 |
498 |
|
475 |
1,035 |
|
50.38538146 |
100.7564663 |
|
Conserved hypothetical |
|
XM_002847992.1 |
1,983 |
|
474 |
1,167 |
|
12.62687588 |
28.53054465 |
|
Alkaline phosphatase 4 |
|
XM_002846727.1 |
1,311 |
|
473 |
1,539 |
|
19.05893957 |
56.91121385 |
|
Aspartate |
|
XM_002846830.1 |
1,136 |
|
466 |
1,855 |
|
21.66944905 |
79.16395863 |
|
Conserved hypothetical |
|
XM_002850401.1 |
1,344 |
|
464 |
1,331 |
|
18.23723506 |
48.01099509 |
|
2-oxoisovalerate |
|
XM_002848773.1 |
1,752 |
|
424 |
1,943 |
|
12.78415533 |
53.76511999 |
|
Phosphate-repressible |
|
XM_002846918.1 |
729 |
|
411 |
4,370 |
|
29.78204926 |
290.6135297 |
|
Conserved hypothetical |
|
XM_002851113.1 |
828 |
|
390 |
1,142 |
|
24.88138517 |
66.86482121 |
|
PEP phosphonomutase |
|
XM_002848106.1 |
1,056 |
|
336 |
5,233 |
|
16.80798467 |
240.2418907 |
|
Lysine amidinotransferase |
|
XM_002848786.1 |
4,317 |
|
289 |
1,080 |
|
3.536356813 |
12.12840323 |
|
Enniatin synthetase |
|
XM_002842695.1 |
897 |
|
276 |
1,082 |
|
16.25387528 |
58.47856922 |
|
Conserved hypothetical |
|
XM_002845832.1 |
573 |
|
210 |
2,262 |
|
19.35998234 |
191.3814758 |
|
Uncharacterized protein |
|
XM_002842669.1 |
696 |
|
208 |
498 |
|
15.7868099 |
34.6882207 |
|
Uncharacterized protein |
|
XM_002848105.1 |
1,566 |
|
193 |
1,203 |
|
6.510372459 |
37.24223963 |
|
Conserved hypothetical |
|
XM_002850196.1 |
1,227 |
|
143 |
780 |
|
6.156469876 |
30.8185329 |
|
Conserved hypothetical |
|
XM_002846540.1 |
1,068 |
|
105 |
431 |
|
5.193478409 |
19.56446327 |
|
Alpha/beta hydrolase |
3. ITCZ-induced increase in gene expression
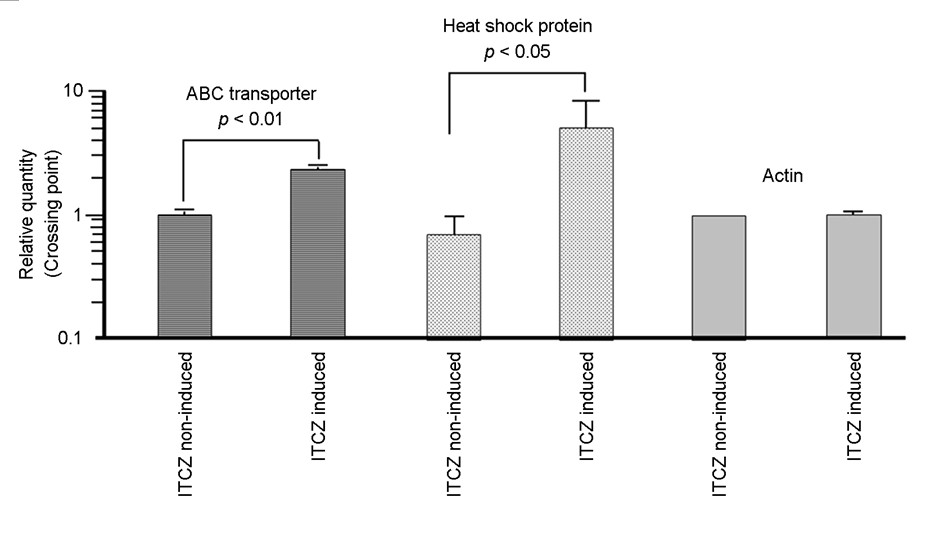
Transcript levels of the heat shock protein and ABC trans- porter genes in ITCZ-tolerant M. canis strain DTMU25-001 were compared between samples cultured in SDB containing 0.2 μg/mL ITCZ versus ITCZ-free conditions. The results of RT-qPCR analyses confirmed that ITCZ stimulation increased the expression of the ABC transporter gene approximately 2-fold and the heat shock protein gene 6-fold in M. canis cells compared with the unstimulated condition (Fig. 1).
4. Data summary
Updated sequences were submitted to GenBank under the following accession numbers: for data of the transcriptome analysis of ITCZ-induced/non-induced strain DTMU25-001, BioProject: PRJDB35662, and Run: DRR706743-DRR706744.
To the best of our knowledge, this is the first report of a transcriptome analysis conducted to clarify the ITCZ tolerance mechanism of an ITCZ-tolerant M. canis strain from human dermatophytosis. The MICs and MFCs for this strain suggest that it is particularly tolerant to ITCZ. Therefore, a trans- criptome analysis was performed to analyze gene expression in samples cultured in the presence and absence of ITCZ. To investigate the adaptive transcriptional responses in this experiment, RNA was extracted after culturing at an ITCZ concentration of 0.2 μg/mL, following Martins et al., who used a concentration of 0.14 μg/mL TBF to evaluate gene expression in four species of Trichophyton12. Although TBF and ITCZ use different chemicals, this concentration was chosen to examine the response to the antifungal agent while avoiding cell death. The transcriptome analysis indicated that the expression levels of the heat shock protein gene (GenBank accession no. XM_002843435.1) and ABC transporter gene (GenBank accession no. XM_002847376.1) increased the most following culture in the presence of ITCZ (Supplemental Table 1). The results of RT-qPCR analyses confirmed that ITCZ stimulation increased the expression of the ABC transporter gene approximately 2-fold and that of the heat shock protein gene 6-fold in M. canis cells over the unstimulated condition (Fig. 1). The RT-qPCR analysis thus corroborated the results of the transcriptome analysis. However, because the RT-qPCR results only pertain to two gene variations per strain, they merely serve to confirm the findings of this transcriptome analysis.
The heat shock protein and ABC transporter are reportedly involved in the pathogenicity and antifungal drug resistance of anthropophilic dermatophytes8,13. For example, different HSPs were detected during the infection and invasion of the host tissue, such as the overexpression of Hsp 30, HSP 60, HSP 70, HSP 78, and HSP 70 and of HSP 90, HSP-related gene hsf1, and HSPSSc1 in T. rubrum8,13. Resistance to azoles has also been repeatedly reported and is associated with the overexpression of two genes (MDR2 and MDR3) encoding a multidrug transporter of the ABC transporter genes, giving rise to multidrug efflux outside the cell in T. rubrum8. In a recent study, we conducted a transcriptome analysis under the same conditions to clarify the multi-azole-resistance mechanism of a strain of M. canis isolated from a feline der- matophytosis14. The results of that experiment also showed increased expression of the same two genes. Moreover, we investigated whether sensitivity to azoles increased during cotreatment with milbemycin, which inhibits the ABC trans- porter in dermatophytes. The milbemycin counteracted the ITCZ and RVCZ tolerance of the M. canis isolate, suggesting that the strain acquired resistance through the high expression of the ABC transporter14. However, we have not investigated the heat shock protein for the azole-tolerant strains. Therefore, in future studies, we plan to generate heat shock protein and ABC transporter gene disruption in M. canis strains to investigate how these changes affect susceptibility to ITCZ. We would also like to investigate other genes that exhibited increased expression.
Additionally, 22 upregulated genes were identified; how- ever, based on the gene names registered in GenBank, it was unclear whether they were associated with ITCZ tolerance (Supplemental Table 1). Because it would be extremely difficult to create knockout strains for all of these genes, we plan to perform transcriptomic analysis on several ITCZ-tolerant strains and investigate the common upregulated genes.
In contrast, ITCZ stimulation decreased the expression of more genes than it increased. As reported by Carmo et al., Trichophyton interdigitale, which became resistant to ITCZ, ex- hibited a decrease in cellular metabolism15, which is thought to diminish the toxicity of ITCZ and is suspected to be related to a diversion of energy to sustain the high activity of ABC transporters. The results of our transcriptome analysis suggest that cellular metabolic activity was attenuated, which may contribute to the mechanism of ITCZ tolerance in a broad sense. Our study suggests that multiple intracellular functions are involved in the mechanism of tolerance to ITCZ in M. canis. However, further research is needed to fully elucidate the mechanism of resistance.
References
1. Moriello KA, Coyner K. Dermatophytosis. In: Sykes JE, Rankin SC, Papich MG, Weese JS, Little SE editors. Greene's Infectious Diseases of the Dog and Cat. 5th edition. London: Elsevier, 2023:961-967
2. Moriello KA, Coyner K, Paterson S, Mignon B. Diagnosis and treatment of dermatophytosis in dogs and cats: Clinical consensus guidelines of the world association for veterinary dermatology. Vet Dermatol 2017;28:266-e68
Google Scholar
3. Lockhart SR, Chowdhary A, Gold JA. The rapid emer- gence of antifungal-resistant human-pathogenic fungi. Nat Rev Microbiol 2023;21:818-832
Google Scholar
4. Gold JAW, Lipner SR. The rise of antifungal-resistant dermatophyte infections: What dermatologists need to know. Cutis 2025;115:151-154
Google Scholar
5. Hsiao YH, Chen C, Han HS, Kano R. The first report of terbinafine resistance Microsporum canis from a cat. J Vet Med Sci 2018;80:898-900
Google Scholar
6. Kano R, Hsiao YH, Han HS, Chen C, Hasegawa A, Kamata H. Resistance mechanism in a terbinafine-resistant strain of Microsporum canis. Mycopathologia 2018;183:623-627
Google Scholar
7. Kakurai M, Harada K, Kano R, Demitsu T. A case of tinea faciei caused by itraconazole-resistant Microsporum canis: The first report in a human in Japan. J Dermatol 2025;52:e1066-e1067
Google Scholar
8. Dubljanin E, Zunic J, Vujcic I, Colovic Calovski I, Sipetic Grujicic S, Mijatovic S, et al. Host-pathogen interaction and resistance mechanisms in dermatophytes. Pathogens 2024;13:657
Google Scholar
9. CLSI document M38-A2. Reference method for broth dilution antifungal susceptibility testing of filamentous fungi; approved standard: 2nd ed. Philadelphia: Clinical Laboratory Standards Institute, 2008
10. Itoi S, Kano R, Hasegawa A, Kamata H. In vitro activities of antifungal agents against clinical isolates of dermato- phytes from animals. J Vet Med Sci 2012;74:1067-1069
Google Scholar
11. Hiruma J, Nojyo H, Harada K, Kano R. Development of treatment strategies by comparing the minimum in- hibitory concentrations and minimum fungicidal con- centrations of azole drugs in dermatophytes. J Dermatol 2024;51:1515-1518
Google Scholar
12. Martins MP, Franceschini ACC, Jacob TR, Rossi A, Martinez-Rossi NM. Compensatory expression of multidrug-resistance genes encoding ABC transporters in dermatophytes. J Med Microbiol 2016;65:605-610
Google Scholar
13. Sardana K, Sharath S, Khurana A, Ghosh S. An update on the myriad antifungal resistance mechanisms in dermato- phytes and the place of experimental and existential therapeutic agents for Trichophyton complex implicated in tinea corporis and cruris. Expert Rev Anti Infect Ther 2023;21:971-977
Google Scholar
14. Tsurumi K, Oshima Y, Murayama N, Makimura K, Kano R. Transcriptome analysis of multi-azole-resistant Micro- sporum canis isolated from a case of feline dermato- phytosis. J Vet Med Sci 2026;88:26-29
Google Scholar
15. Carmo PHF, Costa MC, Leocádio VAT, Gouveia-Eufrásio L, Emídio ECP, Pimentel SP, et al. Exposure to itraconazole influences the susceptibility to antifungals, physiology, and virulence of Trichophyton interdigitale. Med Mycol 2022;60:myac088
Google Scholar
Congratulatory MessageClick here!
