pISSN : 3058-423X eISSN: 3058-4302
Open Access, Peer-reviewed
pISSN : 3058-423X eISSN: 3058-4302
Open Access, Peer-reviewed

Young Bok Lee
10.17966/JMI.2026.31.2.51 Epub 2026 June 30
Abstract
The emergence of antifungal-resistant dermatophytes worldwide, particularly Trichophyton indotineae, has transformed the clinical and epidemiological landscape of dermatophytosis. Increasing terbinafine (TRB) resistance has raised concerns over treatment failure, relapse, and the occurrence of chronic infection. To review the current knowledge of TRB-resistant dermatophytosis and onychomycosis, focusing on resistance mechanisms, epidemiology, diagnosis, and treatment. TRB resistance is mainly mediated by point mutations in the squalene epoxidase (SQLE) gene, with amino acid substitutions, such as L393F, F397L, and H440Y, conferring high minimum inhibitory concentrations. Additional mechanisms, such as efflux pump overexpression, CYP51B gene amplification, and biofilm formation, contribute to multidrug resistance and clinical recalcitrance. Trichophyton indotineae has emerged as the dominant species in India and is currently spreading globally. Onychomycosis acts as a critical reservoir for resistant strains because of subtherapeutic drug concentrations, high fungicidal thresholds, and biofilm-mediated protection. Diagnostic confirmation using SQLE sequencing and agar-based screening is essential, given that clinical breakpoints have not yet been defined. Itraconazole is the most effective systemic alternative for TRB-resistant infections, with newer azoles and topical agents used as adjuncts. The increasing prevalence of TRB-resistant dermatophytes underscores the need for enhanced molecular surveillance, judicious antifungal use, and restriction of corticosteroid-antifungal combination misuse. Integrated diagnostic and therapeutic approaches are essential for controlling the global spread of resistant Trichophyton species.
Keywords
Dermatophyte Drug resistance Onychomycosis Squalene epoxidase Terbinafine
Dermatophytoses, or tinea, are superficial fungal infections affecting keratinized tissues (skin, hair, and nails)1. These infections, affecting approximately 20-25% of the global population2, are primarily caused by dermatophytes of the genus Trichophyton, with T. rubrum traditionally representing the most common causative agent3. Although historically considered easily treatable, the emergence of recalcitrant infections, commonly characterized by extensive and difficult-to-treat tinea corporis and cruris, has become a global con- cern, stimulating renewed research interest1. Infections caused by T. interdigitale and T. mentagrophytes have significantly increased in some regions4.
Terbinafine (TRB), an allylamine inhibitor of squalene epoxi- dase (SQLE), remains the first-line systemic agent for dermato- phytoses, including onychomycosis, owing to its fungicidal effect, favorable pharmacokinetic profile, and high keratin adherence5. However, the increased documentation of anti- fungal resistance over the past decade significantly challenges patient management and clinical success, which also increases antibiotic resistance1.
This study is a narrative, descriptive review of the current literature on TRB resistance in dermatophytes, which particu- larly focus on onychomycosis as a clinically relevant reservoir of resistant strains. Relevant publications were identified through a broad literature search in PubMed and related databases using the following keywords: "terbinafine re- sistance," "onychomycosis," and "dermatophytosis." Original research articles, epidemiological studies, and clinically relevant reviews were included for a comprehensive overview of resistance mechanisms, epidemiology, diagnostic challenges, and treatment considerations, whereas studies not directly related to antifungal resistance or clinical management were excluded. This narrative review includes literature published between 2003 and 2024, starting from the first confirmed clinical report of TRB-resistant dermatophytes to recent studies analyzing the global emergence and dissemination of resist- ant Trichophyton species, including Trichophyton indotineae. References were chosen based on clinical relevance, empha- sizing studies reporting in vitro susceptibility data, molecular resistance mechanisms, and epidemiological trends.
Antifungal resistance in dermatophytes was rarely reported until the mid-2010s but has increased rapidly since 2017, coinciding with a notable increase in recalcitrant dermato- phytosis6,7. This epidemiological shift is primarily linked to the emergence and spread of TRB-resistant strains and is increasingly recognized as an emerging international clinical and public health concern (Table 1). The identification of Trichophyton indotineae, formerly classified within the T. mentagrophytes/T. interdigitale complex, is particularly alarm- ing because of its high TRB resistance levels and enhanced transmissibility8.
|
Region/ |
Predominant |
Period of |
Estimated prevalence of |
Notable |
|
India8-12 |
T. indotineae |
2017-2019 |
52-94% of dermatophyte |
Epidemic of chronic/recalcitrant |
|
Iran13 |
T. indotineae |
2008-2010 |
Endemic presence; multiple |
One of earliest non-Indian countries with endemic |
|
Iraq13 |
T. indotineae |
2019 |
39% of T.
mentagrophytes |
High prevalence confirmed |
|
Japan/Vietnam/ |
T. indotineae |
2015-2020 |
Isolated cases |
Spread through regional |
|
Switzerland17 |
T. rubrum, |
2013-2021 |
TRB resistance increased |
Nationwide surveillance |
|
Denmark12,18 |
T. mentagrophytes/ |
2019-2020 |
56-61% of isolates resistant |
High prevalence, mainly imported cases |
|
Germany/France/ |
T. indotineae, |
2019-2023 |
Sporadic to moderate |
Most cases linked to travel |
|
Canada19 |
T. indotineae |
2020s |
Detected in patients with |
First confirmed |
|
USA20 |
T. indotineae |
2020s |
Case reports only |
Imported infections, limited |
|
Australia21 |
T. indotineae |
2021 |
Sporadic |
Imported cases, confirmed |
|
TRB, terbinafine; ITS, internal transcribed spacer |
||||
India is the epicenter of this phenomenon. Since 2017, T. indotineae has largely replaced T. rubrum as the main cause of tinea cruris, corporis, and faciei in multiple regions8. Molecular epidemiological studies conducted between 2017 and 2019 reported that T. indotineae accounts for approxi- mately 52-94% of dermatophyte isolates, with the highest prevalence reported in North and West India8-10. The overall frequency of TRB resistance among T. indotineae isolates is approximately 75% nationwide, although substantial regional variation exists, ranging from 16% in South India to 77% in West India8,11,12.
Beyond India, TRB resistance have also been increasingly reported across Asia6. Endemic circulation has been suggested in Iran, with retrospective detection dating back to 2008-2010, whereas recent studies in Iraq demonstrate that T. indotineae constitutes a substantial proportion of clinical isolates13. Additional cases have also been reported in Japan, Vietnam, and Bangladesh14,15.
In Europe, TRB-resistant Trichophyton infections have been reported with increasing frequency, commonly associated with travel to endemic regions16. The total proportion of TRB-resistant T. rubrum and T. mentagrophytes/T. interdigitale strains in Switzerland increased from 0.63% in 2013 to 1.3% in 202117. Increasing resistance has been reported in Danish Trichophyton isolates, comprising 56% and 61% of samples reviewed in 2019-202012,18. Additional cases have been reported in Germany, France, Belgium, Poland, and Greece12.
Outside Europe, TRB-resistant T. indotineae strains have been isolated in patients with superficial dermatophyte infec- tions in Canada19, the USA20, and Australia21, highlighting the ongoing international dissemination of this resistant species.
Antifungal resistance of fungi can be categorized as innate (primary) or acquired (secondary)22. Acquired resistance develops over time, often due to drug exposure. Its underlying mechanisms are varied and complex (Fig. 1)12.
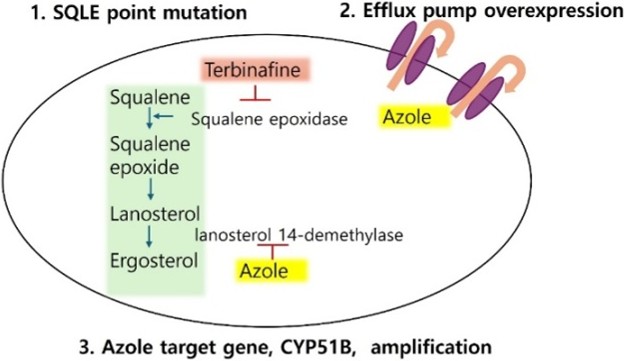
1. SQLE point mutations as the primary mechanism of TRB resistance in dermatophytes
TRB resistance in dermatophytes is predominantly mediated by nonsynonymous point mutations in the gene-encoding SQLE 4, the molecular target of allylamine antifungals17. These mutations result in amino acid substitutions that alter the TRB-binding pocket of the SQLE enzyme, thereby reducing drug affinity and antifungal susceptibility23.
Studies reported that several recurrent SQLE substitutions have been consistently identified in TRB-resistant Trichophyton isolates. The most frequently reported mutations include substitutions at Leu393 (L393F, L393S), Phe397 (F397L, F397I, F397V), Phe415 (F415S, F415I, F415V), and His440 (H440Y). Among these, F397L is the most prevalent and is particularly dominant in resistant strains belonging to the Trichophyton mentagrophytes/Trichophyton indotineae complex6,13,17,24-29.
Functional and susceptibility studies showed that L393F, L393S, and F397L substitutions are generally associated with markedly elevated minimum inhibitory concentrations (MICs) to TRB, with F397L and L393F conferring the highest resistance levels17,25-27. Conversely, Phe415 and His440 mutations are observed less frequently and show more variable effects on MICs in T. rubrum and T. mentagrophytes isolates6,17,24,25,27,29.
The clinical relevance of SQLE mutations is supported by their strong association with treatment failure in patients receiving TRB therapy3,17,26-28. Collectively, these findings estab- lish SQLE point mutations as the main molecular determinant of TRB resistance in dermatophytes. Although amplification of the salicylate 1-monooxygenase (salA) gene has been reported as additional mechanism in T. rubrum, it plays a secondary role compared with SQLE alterations30.
In contrast to TRB resistance, azole resistance in dermato- phytes is most frequently mediated by overexpression of drug efflux pumps that actively lower intracellular antifungal concentrations. These transporters, belonging mainly to ATP-binding cassette transporters (TruMDR1, TruMDR2, TruMDR3, TruMDR5) and major facilitator superfamily transporters (TruMFS1, TruMFS2), actively pump the antifungal drugs out of the fungal cell25,31-35. TruMDR3 overexpression confers to voriconazole resistance and contributes to itraconazole resist- ance. Its disruption in resistant T. rubrum strains eliminates voriconazole resistance and partially reduces itraconazole resistance, indicating its critical function in azole efflux31,36. TruMDR2 is also upregulated in resistant strains and is a major contributor to itraconazole resistance, TruMDR2 suppression eliminates itraconazole resistance even in the absence of TruMDR3, confirming its dominant contribution to this pheno- type35,36.
Major facilitator superfamily transporters, particularly Tru- MFS1 and TruMFS2, are also implicated in azole resistance, which is upregulated in multidrug-resistant isolates31,37. The geographic distribution of these resistances is expanding, with molecular surveillance studies confirming their presence in resistant Trichophyton isolates from multiple continents25,31-35.
1. CYP51B gene amplification as a novel mechanism of azole resistance in T. indotineae
Beyond efflux pump overexpression, CYP51B gene amplifi- cation has recently emerged as a distinct and clinically relevant mechanism of azole resistance in T. indotineae. This mech- anism results in overexpression of the target enzyme lanosterol 14-demethylase, thereby diminishing azole efficacy without requiring target site mutations in Erg11, a well-characterized resistance mechanism in yeasts25,33. In T. indotineae, increased copy number of CYP51B elevated enzyme levels, allowing the fungus to maintain ergosterol biosynthesis despite azole exposure. This mechanism is distinct from point mutations in ERG11 (CYP51) that confer resistance in Candida and other yeasts and highlights a unique adaptation in dermatophytes.
Functional studies demonstrate that CYP51B downregula- tion by RNA interference restores azole susceptibility, whereas overexpression confers resistance, confirming the causal role of gene amplification33. This finding broadens the spectrum of known resistance mechanisms in dermatophytes beyond efflux pump overexpression and, rarely, target site mutations. The clinical implication is that molecular diagnostics for azole resistance in Trichophyton should include assessment of the CYP51B copy number, not just sequence analysis for point mutations.
2. Nonmutational mechanisms of reduced antifungal susceptibility
Dermatophytes can form biofilms, notably observed as dense white fungal masses (dermatophytomas) in infected nails38. The physical and metabolic properties of biofilms protect fungi from antimicrobials, thereby promoting drug resistance, particularly in onychomycosis12. Dermatophytes can also secrete proteins (heat shock proteins) in response to drug exposure stress, which stabilizes the cell and may facilitate the occurrence of resistance mechanisms over time38,39.
Onychomycosis is a critical clinical and biological niche for the emergence, selection, and persistence of TRB-resistant dermatophytes. It is the most frequent clinical presentation among patients with clinically relevant TRB resistance (13 out of 22 cases in one Swiss cohort)17. Historically, TRB resistance was first documented in a T. rubrum isolate sourced from a single patient diagnosed with onychomycosis who failed prolonged TRB treatment (24 weeks)40.
The nail bed and nail plate are considered a likely cradle for the development of antifungal resistance, particularly to TRB, because drug concentrations achieved in the nail are substantially lower than those in the stratum corneum, often falling below the MIC required to inhibit dermatophyte growth. This subtherapeutic exposure is compounded by the fact that experimental models show that the minimum fungicidal concentration (MFC) required to eradicate fungi in nail tissue is much higher than the MIC determined in standard assays, reflecting the unique physical and biochemical barriers of the nail environment41,42.
Additionally, biofilm formation by dermatophytes within the nail creates microenvironments where fungi are exposed to subinhibitory drug concentrations, further promoting the selection of resistant mutants. Biofilms impede drug pene- tration and facilitate the survival of persister cells, which can acquire resistance through stress-induced genetic and epi- genetic changes41,43. Chronic or incomplete exposure to TRB, especially in the context of prolonged or intermittent therapy, increases the risk of resistance development, as demonstrated by the emergence of strains with high MICs and MFCs during therapy41.
These factors collectively make the nail unit a reservoir for resistant dermatophytes, with clinical studies documenting the selection and persistence of TRB-resistant strains in onycho- mycosis patients who fail therapy41,42. This underscores the importance of antifungal stewardship, accurate diagnosis, and consideration of alternative or combination therapies in recalcitrant cases42.
The rise in TRB-resistant onychomycosis is clinically signifi- cant because it is a major contributor to increasing reports of recalcitrant dermatophytosis across multiple regions42. Onychomycosis acts as a reservoir for resistant Trichophyton species, potentiating antifungal resistance through several mechanisms; low drug concentrations in the nail, high fungi- cidal requirements, and biofilm formation, which exposes fungi to subinhibitory drug levels and promotes selection of resistant mutants42.
This resistance leads to persistent and atypical clinical pre- sentations, such as extensive, noninflammatory, or treatment-refractory tinea, and increases the risk of transmission to other body sites and individuals44. The emergence of multidrug-resistant strains, especially Trichophyton indotineae and T. rubrum, complicates management and often results in high relapse rates and therapeutic failure with standard regimens44.
The rising prevalence of TRB-resistant onychomycosis, particularly due to Trichophyton rubrum and T. indotineae, has led to increased rates of treatment failure and a growing need for alternative therapies7,45-47. These resistant infections are now documented globally and are associated with high rates of relapse and chronicity, especially in the context of nail disease where drug penetration is limited and biofilm formation is common46,47.
Diagnostic confirmation is essential before changing therapy. Mycological identification, antifungal susceptibility testing, and, where available, molecular detection of resistance muta- tions (SQLE gene mutations) are recommended to guide management and support antifungal stewardship7,48. This approach helps avoid unnecessary exposure to second-line agents and limits the development of further resistance.
In patients with onychomycosis caused by TRB-resistant strains, switching from TRB to a systemic azole, most com- monly itraconazole, is a commonly adopted therapeutic strategy supported by available clinical evidence7,46,48. Griseo- fulvin and fluconazole have limited efficacy against dermato- phyte onychomycosis and are therefore less favored in cases of TRB resistance7,46. Among systemic azoles, itraconazole is the most widely used second-line agent in clinical practice for TRB-resistant onychomycosis. The most widely used and evidence-based dosing regimens are continuous daily dosing (200 mg once daily for 8-12 weeks) and pulse dosing (400 mg /day for 1 week per month, repeated for 3-6 months)49.
Alternative or adjunctive options include newer-generation azoles such as posaconazole, voriconazole, fosravuconazole, and oteseconazole. Although these agents have shown efficacy in recalcitrant cases, they are not FDA-approved for onychomycosis and are generally reserved for patients who fail both TRB and itraconazole49. Topical agents (efinaconazole, ciclopirox, tavaborole) are effective against both susceptible and resistant strains and may be used in combination with oral therapy to enhance cure rates, particularly in cases with limited nail involvement or as adjuncts to systemic therapy42. Treatment selection should be individualized based on clinical severity, antifungal susceptibility results, drug availability, and patient-specific factors, in the absence of universally accepted treatment guidelines for TRB-resistant onychomycosis.
Strategies to reduce the emergence and spread of TRB-resistant dermatophytes are summarized in Table 2. Reducing the spread and emergence of TRB-resistant strains requires a multi-pronged approach addressing pharmaceutical use, patient behavior, and diagnostic practices.
|
Strategy |
Key measures |
Rationale/mechanism |
Expected outcome |
|
Control topical |
Prohibit or restrict over-the-counter Enforce prescription-only regulations |
Potent topical steroids suppress local immune defense
and mask symptoms, promoting deeper fungal invasion and persistence under
subtherapeutic antifungal exposure. |
Reduced misuse-related recalcitrant tinea and slower
emergence of resistant Trichophyton
strains. |
|
Optimize oral |
Avoid sub-MIC dosing regimens. Ensure adherence to complete Avoid inappropriate co-use of topical
corticosteroids during systemic therapy. |
Sub-MIC exposure enables survival and adaptive
selection of mutant strains; incomplete courses provide a reservoir |
Decreased selection pressure and lower recurrence
rates of |
|
Implement |
Introduce rapid diagnostic tools, such Use results to promptly switch therapy (e.g., from
terbinafine to itraconazole). |
Early molecular identification of SQLE mutations enables class switch
before resistance amplification or transmission. |
Timely, effective management and reduced spread of
resistant strains. |
|
TRB, terbinafine; MIC, minimum inhibitory
concentration; PCR, polymerase chain reaction; SQLE, squalene epoxidase |
|||
1. Control topical medication misuse
Unsupervised topical use of fixed-dose combination creams containing potent topical corticosteroids such as clobetasol propionate combined with antifungal agents is a major contributor to treatment failure, resistance, and recalcitrant dermatophytosis. Potent topical corticosteroids suppress local immune responses and may mask clinical symptoms. This suppression can promote deeper fungal invasion and reduce the effectiveness of antifungal therapy, particularly when the antifungal component is present at subtherapeutic concen- trations50,51. These observations have led to growing concern regarding the widespread availability and inappropriate use of such combination products in affected regions51.
2. Optimize oral antifungal usage
Subminimum inhibitory concentration (sub-MIC) exposure resulting from inadequate dosing or premature discontinuation of antifungal therapy has been implicated in the selection of resistant fungal populations. Ensuring that patients complete the full prescribed treatment course is equally important, particularly when topical antifungal–corticosteroid combin- ation therapy has been used. Sub-MIC exposure allows fungal pathogens to survive and adapt, selecting for resistant mutants and facilitating the emergence of high-level resistance, as demonstrated across multiple infectious disease models, including fungi. Incomplete or inadequate dosing fails to eradicate the pathogen, providing a selective environment for resistant strains to proliferate52.
3. Targeted therapy
Where available, rapid diagnostic approaches, including PCR-based detection and sequencing of SQLE mutations, may facilitate earlier confirmation of TRB resistance. Timely identification of resistance can support informed therapeutic decision-making, including consideration of alternative anti- fungal classes53,54.
Routine antifungal susceptibility testing for dermatophytes remains limited in clinical practice due to technical, methodo- logical, and interpretative challenges, including the lack of standardized protocols, slow fungal growth, and practical constraints in clinical laboratories. First, universally accepted and standardized in vitro susceptibility testing protocols for dermatophytes are lacking. Although recent advances, such as the EUCAST E.Def 11.0 method, have improved repro- ducibility, these assays remain technically demanding and are not widely implemented in routine clinical laboratories55,56. Second, dermatophytes exhibit slow growth and poor sporu- lation, which prolong incubation and complicate inoculum preparation. As a result, turnaround times commonly extend to 7-14 days57,58.
Standard culture methods are further limited by high false-negative rates and poor discrimination between closely related species, such as Trichophyton indotineae, T. mentagrophytes, and T. interdigitale. Consequently, molecular methods are often required for accurate species identification and resist- ance detection44. In addition, antifungal susceptibility testing is labor-intensive, requires specialized equipment and expertise, and incurs substantial costs. These factors limit its feasibility in routine clinical practice58.
Emerging diagnostic tools for detecting TRB resistance in dermatophytes include SQLE gene sequencing, agar-based screening using Sabouraud dextrose agar with TRB, molecular assays such as PCR and sequencing analysis, and MALDI-TOF mass spectrometry for species differentiation. Compared with potassium hydroxide examination and conventional culture, molecular methods such as PCR and sequencing offer higher sensitivity and species-level specificity, while agar-based screening provides a pragmatic phenotypic approach for rapid resistance detection in routine clinical practice.
SQLE gene sequencing is considered the gold standard for confirming TRB resistance, as it directly identifies resistance-associated mutations17. Agar-based screening using Sabou- raud dextrose agar containing 0.2 μg/mL TRB provides a rapid, sporulation-independent phenotypic screening method. Isolates that grow in the presence of TRB are highly likely to harbor SQLE mutations17,56. Molecular assays, such as real-time PCR and ARMS-PCR, enable rapid detection of com- mon SQLE mutations, but may miss novel or rare variants; sequencing analysis remains necessary for comprehensive mutation profiling53. MALDI-TOF mass spectrometry is increas- ingly used for accurate species identification, which is critical for distinguishing T. indotineae from T. mentagrophytes and T. interdigitale, as these species differ in resistance profiles and clinical management59.
Because standardized clinical breakpoints for dermatophytes have not yet been established, interpretation of culture-based susceptibility results relies on MIC distributions and epidemio- logical cut-off values (ECOFFs/ECVs) to inform antifungal therapy. Tentative MIC thresholds (MIC ≥ 0.5-1 μg/mL for TRB) and upper limits of wild-type MIC distributions help identify non-wild-type (resistant) isolates and inform treat- ment decisions60. ECOFFs distinguish wild-type from non-wild-type populations but do not directly predict clinical out- comes. Therefore, clinical correlation and mutation analysis remain essential for therapeutic decision-making60. In practice, isolates with MICs above the ECOFF or with SQLE mutations should prompt a switch to an alternative antifungal, such as itraconazole60.
In clinical practice, initial diagnosis for dermatophytosis is typically based on KOH examination and culture, followed by agar-based screening or molecular testing in cases of treatment failure or suspected TRB resistance. Sequencing-based confirmation is particularly valuable when therapeutic decisions depend on definitive resistance status. However, in Korea, PCR-based dermatophyte identification and molecular resistance testing are not routinely available, even in tertiary referral centers, and are currently limited to selected institu- tions for research purposes. These molecular assays are not covered by the national health insurance system and are rarely incorporated into routine clinical practice. Consequently, diagnostic workflows in Korea rely predominantly on KOH examination and conventional culture, and treatment decisions for suspected TRB resistance are frequently made empirically, based on clinical response rather than molecular confirmation. This highlights the need for realistic diagnostic approaches that support timely therapeutic decisions in settings where molecular resistance testing is not readily accessible.
The emergence and global spread of TRB-resistant dermato- phytes are a critical shift in the epidemiology of dermato- phytosis and pose an increasing clinical challenge. Addressing this threat requires coordinated surveillance to monitor resistance patterns and species distribution, particularly the expansion of T. indotineae.
Effective containment also depends on antifungal steward- ship, such as judicious use of systemic and topical antifungals, avoidance of subtherapeutic exposure, and regulation of inappropriate corticosteroid-antifungal combinations that drive resistance selection.
Finally, advances in diagnostic innovation are essential. Wider access to molecular identification, SQLE mutation detection, and pragmatic screening approaches will enable timely recognition of resistance and guide appropriate therapy. Together, integrated surveillance, stewardship, and improved diagnostics will be central to mitigating the future impact of TRB-resistant dermatophytosis.
References
1. 1. Hill RC, Caplan AS, Elewski B, Gold JAW, Lockhart SR, Smith DJ, et al. Expert panel review of skin and hair dermatophytoses in an era of antifungal resistance. Am J Clin Dermatol 2024;25:359-389
Google Scholar
2. 2. Chanyachailert P, Leeyaphan C, Bunyaratavej S. Cutane- ous fungal infections caused by dermatophytes and non-dermatophytes: An updated comprehensive review of epidemiology, clinical presentations, and diagnostic testing. J Fungi (Basel) 2023;9:669
Google Scholar
3. 3. Barac A, Stjepanovic M, Krajisnik S, Stevanovic G, Paglietti B, Milosevic B. Dermatophytes: Update on clinical epidemiology and treatment. Mycopathologia 2024;189: 101
Google Scholar
4. 4. Monod M. Antifungal resistance in dermatophytes: Emerging problem and challenge for the medical com- munity. J Mycol Med 2019;29:283-284
Google Scholar
5. 5. Darkes MJ, Scott LJ, Goa KL. Terbinafine: A review of its use in onychomycosis in adults. Am J Clin Dermatol 2003; 4:39-65
Google Scholar
6. 6. Shen JJ, Arendrup MC, Verma S, Saunte DML. The emerging terbinafine-resistant Trichophyton epidemic: What is the role of antifungal susceptibility testing? Dermatology 2022;238:60-79
Google Scholar
7. 7. Gupta AK, Mann A, Polla Ravi S, Wang T. An update on antifungal resistance in dermatophytosis. Expert Opin Pharmacother 2024;25:511-519
Google Scholar
8. 8. Jabet A, Normand AC, Brun S, Dannaoui E, Bachmeyer C, Piarroux R, et al. Trichophyton indotineae, from epidemiology to therapeutic. J Mycol Med 2023;33: 101383
Google Scholar
9. 9. Thakur S, Spruijtenburg B, Abhishek, de Groot T, Meijer EFJ, Narang T, et al. Amplified fragment length polymor- phism genotyping of Trichophyton indotineae indicates possible zoonotic transmission. Med Mycol 2025;63: myaf020
Google Scholar
10. 10. Tang C, Kong X, Ahmed SA, Thakur R, Chowdhary A, Nenoff P, et al. Taxonomy of the Trichophyton menta- grophytes/T. interdigitale species complex harboring the highly virulent, multiresistant genotype T. indotineae. Mycopathologia 2021;186:315-326
Google Scholar
11. 11. Ebert A, Monod M, Salamin K, Burmester A, Uhrlaß S, Wiegand C, et al. Alarming India-wide phenomenon of antifungal resistance in dermatophytes: A multicentre study. Mycoses 2020;63:717-728
Google Scholar
12. 12. Gupta AK, Talukder M, Carviel JL, Cooper EA, Piguet V. Combatting antifungal resistance: Paradigm shift in the diagnosis and management of onychomycosis and dermatomycosis. J Eur Acad Dermatol Venereol 2023; 37:1706-1717
Google Scholar
13. 13. Mahmood HR, Shams-Ghahfarokhi M, Salehi Z, Razzaghi-Abyaneh M. Epidemiological trends, antifungal drug susceptibility and SQLE point mutations in etiologic species of human dermatophytosis in Al-Diwaneyah, Iraq. Sci Rep 2024;14:12669
Google Scholar
14. 14. Ngo TMC, Santona A, Ton Nu PA, Cao LC, Tran Thi G, Do TBT, et al. Detection of terbinafine-resistant Trichophyton indotineae isolates within the Trichophyton mentagro- phytes species complex isolated from patients in Hue City, Vietnam: A comprehensive analysis. Med Mycol 2024;62:myae088
Google Scholar
15. 15. dos Santos AR, Uhrlaß S, Nenoff P, Gold JAW, Bhuiyan MSI, Goturu S, et al. Global emergence of antifungal-resistant dermatophytosis caused by Trichophyton indo- tineae (Formerly T. mentagrophytes ITS genotype VIII): A genomic investigation involving 14 countries. Mycoses 2025;68:e70101
Google Scholar
16. 16. Sacheli R, Hayette MP. Antifungal resistance in dermato- phytes: Genetic considerations, clinical presentations and alternative therapies. J Fungi (Basel) 2021;7:983
Google Scholar
17. 17. Blanchard G, Amarov B, Fratti M, Salamin K, Bontems O, Chang YT, et al. Reliable and rapid identification of terbinafine resistance in dermatophytic nail and skin infections. J Eur Acad Dermatol Venereol 2023;37:2080-2089
Google Scholar
18. 18. Astvad KMT, Hare RK, Jørgensen KM, Saunte DML, Thomsen PK, Arendrup MC. Increasing terbinafine resistance in danish Trichophyton isolates 2019-2020. J Fungi (Basel) 2022;8:150
Google Scholar
19. 19. Posso-De Los Rios CJ, Tadros E, Summerbell RC, Scott JA. Terbinafine resistant Trichophyton indotineae isolated in patients with superficial dermatophyte infection in Canadian patients. J Cutan Med Surg 2022;26:371-376
Google Scholar
20. 20. Chen E, Ghannoum M, Elewski BE. Treatment-resistant tinea corporis, a potential public health issue. Br J Dermatol 2021;184:164-165
Google Scholar
21. 21. Chua KYL, Halliday CL, Mason A, Vogrin S, Knox J, Chen SC. Optimising the detection of Trichophyton indotineae and its prevalence in a large Australian laboratory. Pathology 2025;57:762-766
Google Scholar
22. 22. Perlin DS, Rautemaa-Richardson R, Alastruey-Izquierdo A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect Dis 2017; 17:e383-e392
Google Scholar
23. 23. Khurana A, Sardana K, Chowdhary A. Antifungal resist- ance in dermatophytes: Recent trends and therapeutic implications. Fungal Genet Biol 2019;132:103255
Google Scholar
24. 24. Mahmood HR, Shams-Ghahfarokhi M, Razzaghi-Abyaneh M. Computational analysis of missense mutations in squalene epoxidase associated with terbinafine resistance in clinically reported dermatophytes. Sci Rep 2025;15: 18612
Google Scholar
25. 25. Gupta AK, Wang T, Mann A, Piguet V, Chowdhary A, Bakotic WL. Mechanisms of resistance against allylamine and azole antifungals in Trichophyton: A renewed call for innovative molecular diagnostics in susceptibility testing. PLoS Pathog 2025;21:e1012913
Google Scholar
26. 26. Kong X, Tang C, Singh A, Ahmed SA, Al-Hatmi AMS, Chowdhary A, et al. Antifungal susceptibility and muta- tions in the squalene epoxidase gene in dermatophytes of the Trichophyton mentagrophytes species complex. Antimicrob Agents Chemother 2021;65:e00056-21
Google Scholar
27. 27. Saunte DML, Hare RK, Jørgensen KM, Jørgensen R, Deleuran M, Zachariae CO, et al. Emerging terbinafine resistance in Trichophyton: Clinical characteristics, squa- lene epoxidase gene mutations, and a reliable EUCAST method for detection. Antimicrob Agents Chemother 2019;63:e01126-19
Google Scholar
28. 28. Caplan AS, Todd GC, Zhu Y, Sikora M, Akoh CC, Jakus J, et al. Clinical course, antifungal susceptibility, and genomic sequencing of Trichophyton indotineae. JAMA Dermatol 2024;160:701-709
Google Scholar
29. 29. Gupta AK, Cooper EA, Wang T, Polla Ravi S, Lincoln SA, Piguet V, et al. Detection of squalene epoxidase muta- tions in United States patients with onychomycosis: Implications for management. J Invest Dermatol 2023; 143:2476-2483 e7
Google Scholar
30. 30. Santos HL, Lang EAS, Segato F, Rossi A, Martinez-Rossi NM. Terbinafine resistance conferred by multiple copies of the salicylate 1-monooxygenase gene in Trichophyton rubrum. Med Mycol 2018;56:378-381
Google Scholar
31. 31. Monod M, Feuermann M, Salamin K, Fratti M, Makino M, Alshahni MM, et al. Trichophyton rubrum azole resistance mediated by a new ABC transporter, TruMDR3. Antimicrob Agents Chemother 2019;63:e00863-19
Google Scholar
32. 32. Kano R, Kimura U, Noguchi H, Hiruma M. Clinical isolate of a multi-antifungal-resistant Trichophyton rubrum. Antimicrob Agents Chemother 2022;66:e0239321
Google Scholar
33. 33. Yamada T, Yaguchi T, Maeda M, Alshahni MM, Salamin K, Guenova E, et al. Gene amplification of CYP51B: A new mechanism of resistance to azole compounds in Trichophyton indotineae. Antimicrob Agents Chemother 2022;66:e00059-22
Google Scholar
34. 34. Shao Y, Shao J, de Hoog S, Verweij P, Bai L, Richardson R, et al. Emerging antifungal resistance in Trichophyton mentagrophytes: Insights from susceptibility profiling and genetic mutation analysis. Emerg Microbes Infect 2025;14:2450026
Google Scholar
35. 35. Fachin AL, Ferreira-Nozawa MS, Maccheroni W, Martinez-Rossi NM. Role of the ABC transporter TruMDR2 in ter- binafine, 4-nitroquinoline N-oxide and ethidium bromide susceptibility in Trichophyton rubrum. J Med Microbiol 2006;55:1093-1099
Google Scholar
36. 36. Yamada T, Yaguchi T, Tamura T, Pich C, Salamin K, Feuermann M, et al. Itraconazole resistance of Tricho- phyton rubrum mediated by the ABC transporter TruMDR2. Mycoses 2021;64:936-946
Google Scholar
37. 37. Bhattacharyya A, Sadhasivam S, Sinha M, Gupta S, Saini S, Singh H, et al. Treatment recalcitrant cases of tinea corporis/cruris caused by T. mentagrophytes - interdigitale complex with mutations in ERG11 ERG 3, ERG4, MDR1 MFS genes & SQLE and their potential implications. Int J Dermatol 2023;62:637-648
Google Scholar
38. 38. Koh XQ, Pan JY. Recalcitrant cutaneous fungal infections-A growing problem. Australas J Dermatol 2023;64:315-321
Google Scholar
39. 39. Martinez-Rossi NM, Jacob TR, Sanches PR, Peres NT, Lang EA, Martins MP, et al. Heat shock proteins in dermatophytes: Current advances and perspectives. Curr Genomics 2016;17:99-111
Google Scholar
40. 40. Osborne CS, Leitner I, Hofbauer B, Fielding CA, Favre B, Ryder NS. Biological, biochemical, and molecular char- acterization of a new clinical Trichophyton rubrum isolate resistant to terbinafine. Antimicrob Agents Chemother 2006;50:2234-2236
Google Scholar
41. 41. Mukherjee PK, Leidich SD, Isham N, Leitner I, Ryder NS, Ghannoum MA. Clinical Trichophyton rubrum strain exhibiting primary resistance to terbinafine. Antimicrob Agents Chemother 2003;47:82-86
Google Scholar
42. 42. Gupta AK, Elewski B, Joseph WS, Lipner SR, Daniel CR, Tosti A, et al. Treatment of onychomycosis in an era of antifungal resistance: Role for antifungal stewardship and topical antifungal agents. Mycoses 2024;67:e13683
Google Scholar
43. 43. Martinez-Rossi NM, Bitencourt TA, Peres NTA, Lang EAS, Gomes EV, Quaresemin NR, et al. Dermatophyte resistance to antifungal drugs: Mechanisms and pro- spectus. Front Microbiol 2018;9:1108
Google Scholar
44. 44. Gupta AK, Susmita, Nguyen HC, Liddy A, Talukder M, Wang T, et al. Trichophyton indotineae: Epidemiology, antifungal resistance and antifungal stewardship strat- egies. J Eur Acad Dermatol Venereol 2026;40:29-45
Google Scholar
45. 45. Shaw D, Dogra S, Singh S, Shah S, Narang T, Kaur H, et al. Prolonged treatment of dermatophytosis caused by Trichophyton indotineae with terbinafine or itraconazole impacts better outcomes irrespective of mutation in the squalene epoxidase gene. Mycoses 2024;67:e13778
Google Scholar
46. 46. Song G, Kong X, Li X, Liu W, Liang G. Prior selection of itraconazole in the treatment of recalcitrant Trichophyton indotineae infection: Real-world results from retrospective analysis. Mycoses 2024;67:e13663
Google Scholar
47. 47. Khurana A, Agarwal A, Agrawal D, Panesar S, Ghadlinge M, Sardana K, et al. Effect of different itraconazole dosing regimens on cure rates, treatment duration, safety, and relapse rates in adult patients with tinea corporis/cruris: A randomized clinical trial. JAMA Dermatol 2022;158: 1269-1278
Google Scholar
48. 48. Appelt L, Nenoff P, Uhrlaß S, Krüger C, Kühn P, Eichhorn K, et al. Terbinafine-resistant dermatophytoses and onychomycosis due to Trichophyton rubrum. Hautarzt 2021;72:868-877
Google Scholar
49. 49. Gupta AK, Talukder M, Shemer A, Galili E. Safety and efficacy of new generation azole antifungals in the management of recalcitrant superficial fungal infections and onychomycosis. Expert Rev Anti Infect Ther 2024; 22:399-412
Google Scholar
50. 50. Erbagci Z. Topical therapy for dermatophytoses: Should corticosteroids be included? Am J Clin Dermatol 2004; 5:375-384
Google Scholar
51. 51. Jhaj R, Asati DP, Chaudhary D, Sadasivam B. Topical steroid containing combinations: Burden of adverse effects and why the recent regulatory action may not be enough. Indian J Pharmacol 2021;53:371-376
Google Scholar
52. 52. Drusano GL. Prevention of resistance: A goal for dose selection for antimicrobial agents. Clin Infect Dis 2003; 36:S42-50
Google Scholar
53. 53. Shankarnarayan SA, Shaw D, Sharma A, Chakrabarti A, Dogra S, Kumaran MS, et al. Rapid detection of terbi- nafine resistance in Trichophyton species by amplified refractory mutation system-polymerase chain reaction. Sci Rep 2020;10:1297
Google Scholar
54. 54. Sacheli R, Egrek S, El Moussaoui K, Darfouf R, Adjetey AB, Hayette MP. Evaluation of currently available laboratory methods to detect terbinafine resistant dermatophytes including a gradient strip for terbinafine, EUCAST micro- dilution E.Def 11.0, a commercial real-time PCR assay, squalene epoxidase sequencing and whole genome sequencing. Mycoses 2024;67:e70005
Google Scholar
55. 55. Arendrup MC, Kahlmeter G, Guinea J, Meletiadis J. Subcommittee on Antifungal Susceptibility Testing of The ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). How to: Perform anti- fungal susceptibility testing of microconidia-forming dermatophytes following the new reference EUCAST method E.Def 11.0, exemplified by Trichophyton. Clin Microbiol Infect 2021;27:55-60
Google Scholar
56. 56. Arendrup MC, Jorgensen KM, Guinea J, Lagrou K, Chryssanthou E, Hayette MP, et al. Multicentre validation of a EUCAST method for the antifungal susceptibility testing of microconidia-forming dermatophytes. J Anti- microb Chemother 2020;75:1807-1819
Google Scholar
57. 57. Curatolo R, Juricevic N, Leong C, Bosshard PP. Antifungal susceptibility testing of dermatophytes: Development and evaluation of an optimised broth microdilution method. Mycoses 2021;64:282-291
Google Scholar
58. 58. Markantonatou AM, Samaras K, Zachrou E, Vyzantiadis TA. Comparison of four methods for the in vitro suscepti- bility testing of dermatophytes. Front Microbiol 2020; 11:1593
Google Scholar
59. 59. Cañete-Gibas CF, Mele J, Patterson HP, Sanders CJ, Ferrer D, Garcia V, et al. Terbinafine-resistant dermatophytes and the presence of Trichophyton indotineae in North America. J Clin Microbiol 2023;61:e0056223
Google Scholar
60. 60. Khurana A, Masih A, Chowdhary A, Sardana K, Borker S, Gupta A, et al. Correlation of In vitro susceptibility based on MICs and squalene epoxidase mutations with clinical response to terbinafine in patients with tinea corporis/cruris. Antimicrob Agents Chemother 2018;62: e01038-18
Google Scholar
Congratulatory MessageClick here!
